
سینتیک آنزیم، مکانیسم Michaelis-Menten
در حالی که مطالعه ریاضی واکنشهای شیمیایی بیش از یک قرن است که انجام شده است، تنها اخیراً ابزارهای محاسباتی برای ادغام عددی معادلات سرعت به طور گسترده در دسترس بودهاند. ضرب المثل قدیمی “ضرورت مادر همه اختراعات است” در این مورد صادق است. در اینجا، شما یک تحلیل کلاسیک از یک سیستم واکنش غیر پیش پا افتاده پیدا خواهید کرد و خواهید آموخت که چگونه راه حل ساده شده با راه حل “واقعی” مقایسه می شود.
مولکول های تنبل
پست قبلی وبلاگ مفهوم انرژی فعال سازی را معرفی کرد. این انرژی مورد نیازی است که مولکول ها برای واکنش باید با آن “تأثیر” کنند. برای تعداد زیادی از واکنش های حیاتی، این انرژی بسیار زیاد است. به نظر می رسد برخی از مولکول ها تنبل هستند و کاملاً راضی هستند که در شکل فعلی خود باقی بمانند. این لزوماً چیز بدی نیست – برای مثال گاو را در نظر بگیرید.
گاو، مانند هر ماده آلی، از جمله انسان، ذاتاً از نظر ترمودینامیکی ناپایدار است. آنها علاوه بر محتوای آب، عمدتاً از هیدروکربنهایی تشکیل شدهاند که معمولاً اکسیژندار و نیتراتشده هستند. این یک مخلوط بالقوه قابل احتراق است که به طور قابل توجهی مانند دی اکسید کربن، نیتروژن و آب پایدارتر است. ترمودینامیک نشان می دهد که این شکل بسیار مطلوب تری از نظر انرژی است . خوشبختانه، سینتیک واکنشهای تجزیه آنها بسیار کند است – مولکولها به سادگی انرژی فعالسازی لازم برای تجزیه را ندارند.
حال در نظر بگیرید که یک استیک را روی یک گریل داغ بگذارید. اگر این کار را انجام دهید، خواهید دید که وقتی انرژی فعال سازی به وفور وجود داشته باشد چه اتفاقی می افتد. ابتدا گوشت زغال میکند و در نهایت تودهای از خاکستر (مواد معدنی و غیرآلی گوشت) برای شما باقی میماند. این ما را به نکته بعدی من می رساند: چگونه ما انسان ها از خوردن یک تکه گوشت (به درستی پخته شده) انرژی می گیریم؟ پاسخ از طریق کاتالیزور است .
کاتالیزورها و آنزیم ها
کاتالیزور مادهای است که به نحوی یک مولکول را میکشد و میپیچاند به گونهای که بتواند بر اساس مسیر متفاوت و از نظر انرژی پایینتر واکنش نشان دهد . تأثیر این امر این است که یک واکنش می تواند ناگهان با سرعت قابل توجهی در دماهای بسیار پایین تر انجام شود. کاتالیز در همه جا وجود دارد – حدود 95٪ از تمام محصولات شیمیایی صنعتی حداقل یک مرحله کاتالیزوری را در طول عمر خود طی می کنند.
یک گروه مهم از کاتالیزورها که طی میلیون ها سال برای کار در دماهای بسیار متوسط بهینه شده اند، آنزیم ها هستند . آنزیم ها مولکول های بزرگی هستند (معمولاً از چندین هزار اتم تشکیل شده اند) که به سیستم های بیولوژیکی اجازه می دهند تا واکنش های شیمیایی لازم برای حفظ حیات را انجام دهند. یک تکه نان را برای مدتی بجوید تا شیرین شود – این به دلیل آمیلاز است ، آنزیمی که در بزاق انسان وجود دارد و می تواند نشاسته را به قند تبدیل کند تا هضم راحت تر شود.
پپسین
اگر به مثال استیک خود برگردیم، معلوم میشود که اگر آنزیمها نبود، خود گوشت بسیار سفت، غیراشتهاآور و هضم آن دشوار بود. حتی قبل از اینکه گوشت گاو به بشقاب ما برسد، قصاب گوشت را برای چند هفته آویزان میکند تا به برخی از آنزیمهای طبیعی اجازه دهد تا فرآیند تخریب پروتئین را آغاز کنند – بنابراین گوشت را نرم کرده و طعم کلی آن را افزایش میدهد. در ادامه، آنزیمی که مسئول تخریب پروتئین در دستگاه گوارش ما است، پپسین نام دارد . پپسین در واقع اولین آنزیمی بود که کشف شد، بنابراین من احساس میکنم که نمونه مناسبی برای این وبلاگ است. بیایید نگاهی دقیق تر به نحوه عملکرد پپسین بیندازیم.
در مرحله اولیه، سوبسترای S (در این مورد پروتئین گوشت گاو) باید با آنزیم E مرتبط شود .
E + S → IS
این کمپلکس سوبسترا-آنزیم ES میتواند به خوبی دوباره به زیرلایه و آنزیم جدا شود.
EN → E + S
این دو واکنش معمولاً به صورت یک نوشته میشوند و از توضیح مختصر برای واکنشهای برگشتپذیر استفاده میکنند:
E + S ↔ IS
به جای تفکیک، سوبسترا ممکن است در حالی که به آنزیم کمپلکس می شود واکنش نشان دهد. سپس آنزیم پروتئین محصول را آزاد می کند:
EN → E + P
بنابراین کل سیستم ممکن است به صورت زیر نوشته شود:
E + S ↔ ES → E + P
این واکنش ارتباط نزدیکی با مثال پست قبلی وبلاگ در مورد شیمی دارد . اجازه دهید اکنون سعی کنیم معادلات نرخ این سیستم را فرموله کنیم.
سینتیک Michaelis-Menten
تولید کمپلکس سوبسترا-آنزیم ES به شرح زیر است:
(1)
و مصرف آن:
(2)
ما اکنون به اصطلاح ( شبه ) تقریب حالت پایدار (PSSA یا SSA) را برای ES فرض می کنیم . این تقریب رایج نشان می دهد که یک واسطه واکنشی مانند کمپلکس آنزیم-سوبسترا بسیار واکنش پذیر است. واکنش هایی که گونه های محصول را مصرف می کنند – تفکیک یا واکنش – در مقایسه با تشکیل کمپلکس ذاتاً سریع فرض می شوند. در این مورد می توان تقریب زد که سرعت تغییر غلظت ES ناچیز است. ما مجاز به انجام این کار هستیم حتی اگر غلظت ES ممکن است با ادامه واکنش تغییر کند. آنچه مهم است این است که در مقیاس زمانی واکنشی که ما به آن علاقه داریم، غلظت ES استخیلی سریع به تغییر غلظت کلی سوبسترای واکنش دهنده S پاسخ می دهد .
این تقریب منجر به:
(3)
که معادل است با:
(4)
جمع آوری تمام عبارت های سمت چپ و جمع کردن آنها در یک ثابت تعادلی Michaelis-Menten ، ما بدست می آوریم
(5)
با توجه به غلظت آنزیم موجود در سیستم، غلظت اولیه E را میدانیم و قبلاً یک متغیر برای ES تعریف کردهایم ، بنابراین میتوانیم بنویسیم:
(6)
با وارد کردن این مورد در بالا و حل غلظت ES ، ما باقی میمانیم:
(7)
از این نتیجه می توان برای تعیین میزان تولید پروتئین استفاده کرد:
(8)
جالب است بدانید که در شرایط غلظت بالای سوبسترا (همانطور که معمولاً زمانی که واکنش هنوز زیاد پیش نرفته است) میتوانیم آن را تقریب بزنیم.. تحت این شرایط:
(9)
در این شرایط، میزان مصرف بستر مستقل از خود غلظت بستر است: “رتبه صفر” است. سرعت فقط به غلظت موجود آنزیم بستگی دارد، که واکنش سوبسترا را با حداکثر سرعت ممکن کاتالیز می کند. مقادیرو
با هم برای تعریف سینتیک سیستم واکنش استفاده می شود.
کل این اشتقاق به عنوان مکانیسم Michaelis-Menten شناخته می شود که از لئونور مایکلیس و مود منتن نامگذاری شده است. در بسیاری از واکنش های بیوشیمیایی کاربرد وسیعی پیدا می کند. در زمانی که مایکلیس و منتن نتایج خود را 101 سال پیش منتشر کردند، آنها به کامپیوتر دیجیتالی دسترسی نداشتند که بتواند به سرعت سیستم معادلات دیفرانسیل را برای این چهار گونه ادغام کند. در عوض، با انجام تجزیه و تحلیل فوق با یک تقریب شهودی و با انگیزه فیزیکی، یک رابطه ساده به دست آوردند که به آنها نشان داد غلظت آنزیم و سوبسترا چه تأثیری بر سرعت واکنش آنزیمی دارد. این تقریب چقدر خوب است؟ خوشبختانه، اکنون میتوانیم مدل کامل را محاسبه کنیم – بدون PSSA برای ES – و آن را با نتیجه معادله ساده شده مقایسه کنیم.
سینتیک آنزیم در COMSOL
با استفاده از رابط مهندسی واکنش ، تایپ کردن دو واکنش مرتبط ساده و شهودی است. با استفاده از نمادهای <=>، COMSOL Multiphysics فوراً متوجه میشود که یک واکنش برگشتپذیر باید شبیهسازی شود و زمینههای ورودی لازم را برای ثابتهای سرعت رو به جلو و معکوس (و همچنین ایجاد متغیرهایی برای E، S و ES ) فعال میکند . به همین ترتیب، هنگامی که معادله تولید P در آن تایپ شد، متغیری برای غلظت مربوطه ایجاد می شود. مقادیر k 1 = 1700 M -1 s -1 , k -1 = 10 -3 s -1 و k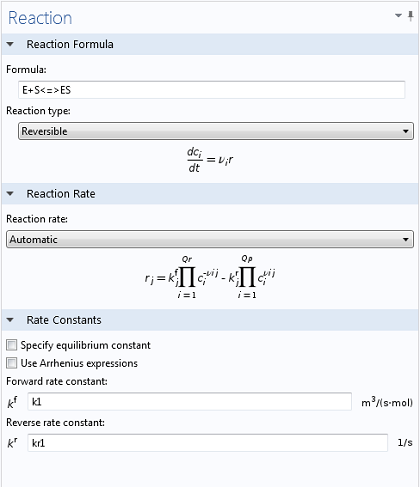
cat = 0.5 s -1 ، و همچنین مقادیر معقول برای غلظت های اولیه E و S ، از ادبیات گرفته شده است ( مشاهده کالریمتری یک واکنش اتصال GroEL-Protein با مشارکت اندک برهمکنش آبگریز ؛ K. Aoki و همکاران J Biol شیمی ج 272، ص 32158-32162، 1997). از آنجایی که ما با چنین تفاوت های قابل توجهی در مقیاس ها سر و کار داریم، دو گونه بسیار متمرکز ( P و S ، شکل 1) به طور جداگانه از گونه های بسیار رقیق نشان داده شده اند ( E و ES ، شکل 2). توجه داشته باشید که این تفاوت در مقیاس ها به طور ذاتی استفاده ما از یک تقریب حالت پایدار را توجیه می کند!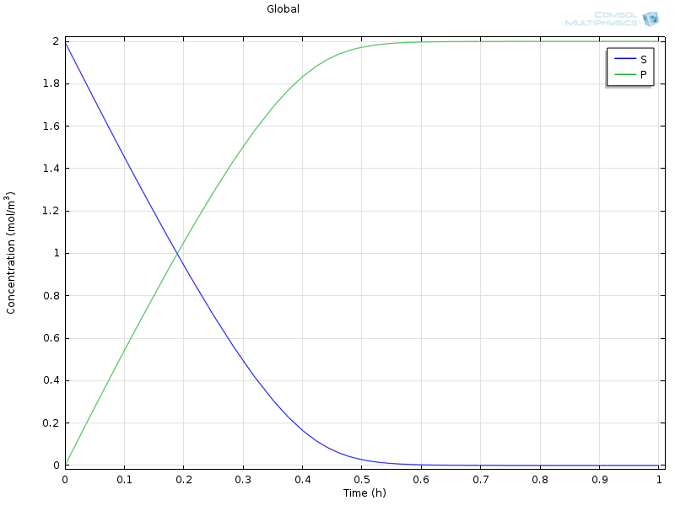
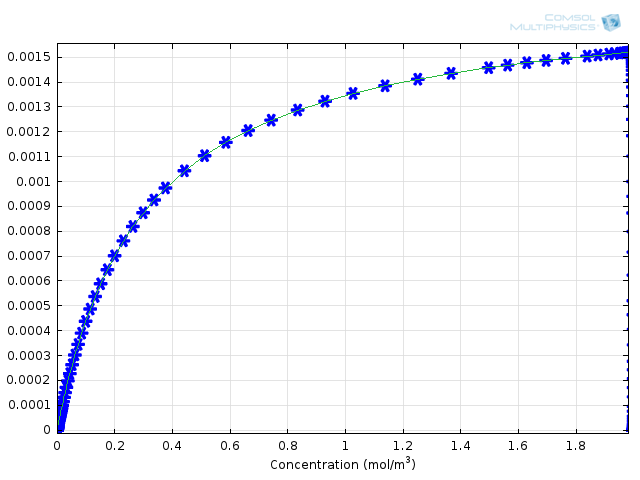
شکل 1: پروفیل های غلظت زیرلایه ( آبی ) و محصول ( سبز ) در طول زمان. شکل 2: غلظت آنزیم ( آبی ) و کمپلکس آنزیم-سوبسترا ( سبز ) در طول زمان. شکل 3: مقایسه سینتیک Michaelis-Menten ( خط جامد سبز ) با مکانیسم واکنش کاملاً یکپارچه ( ستاره های آبی ).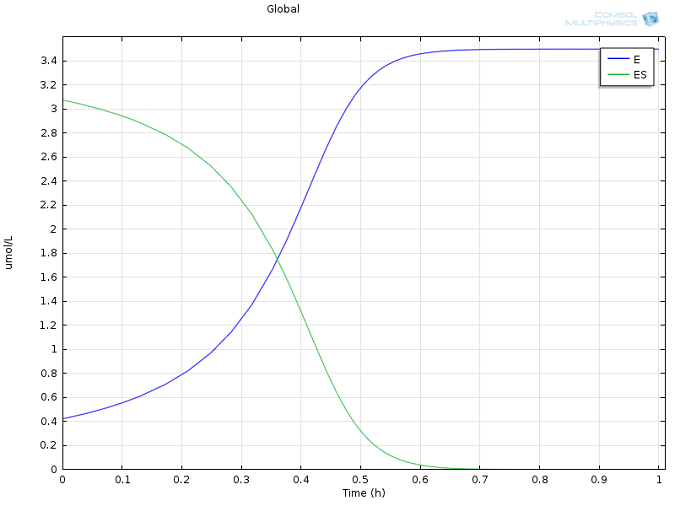
نکات پایانی و مراحل بعدی
چگونه این نتایج می توانند تصوری از خطای مرتبط با مکانیسم Michaelis-Menten به ما ارائه دهند؟ ما می توانیم نتایج را با فرمول محاسبه شده برای سرعت تشکیل محصول مقایسه کنیم. برای انجام این کار، متغیر COMSOL comp1.re.r_2 ، نرخ تولید پروتئین، و kcat*E0*comp1.re.c_S/(Km+comp1.re.c_S) ، عبارت Michaelis-Menten مشتق شده، هستند. بر روی همان گروه پلات 1 بعدی به عنوان توابع غلظت بستر رسم شده است. این شکل 3، بالا را به دست می دهد. می بینید که توافق بین تئوری تقریبی و رویکرد عددی کامل بسیار خوب است.
درست پس از 100 سالگرد مکانیسم Michaelis-Menten، ما به بهترین روشی که می دانیم – با شبیه سازی – از انتشارات پیشگام آنها تقدیر می کنیم.
سایر پست های این مجموعه
- لینک دانلود به صورت پارت های 1 گیگابایتی در فایل های ZIP ارائه شده است.
- در صورتی که به هر دلیل موفق به دانلود فایل مورد نظر نشدید به ما اطلاع دهید.
برای مشاهده لینک دانلود لطفا وارد حساب کاربری خود شوید!
وارد شوید
دیدگاهتان را بنویسید